Methanogens: Methane Producers of the Rumen and Mitigation Strategies
Methanogens are the only known microorganisms capable of methane production, making them of interest when investigating methane abatement strategies. A number of experiments have been conducted to study the methanogen population in the rumen of cattle and sheep, as well as the relationship that methanogens have with other microorganisms. The rumen methanogen species differ depending on diet and geographical location of the host, as does methanogenesis, which can be reduced by modifying dietary composition, or by supplementation of monensin, lipids, organic acids, or plant compounds within the diet. Other methane abatement strategies that have been investigated are defaunation and vaccines. These mitigation methods target the methanogen population of the rumen directly or indirectly, resulting in varying degrees of efficacy. This paper describes the methanogens identified in the rumens of cattle and sheep, as well as a number of methane mitigation strategies that have been effective in vivo.

Introduction
Methane production through enteric fermentation is of concern worldwide for its contribution to the accumulation of greenhouse gases in the atmosphere, as well as its waste of fed energy for the animal. Methane is produced in the rumen and hindgut of animals by a group of Archaea known collectively as methanogens, which belong to the phylum Euryarcheota. Among livestock, methane production is greatest in ruminants, as methanogens are able to produce methane freely through the normal process of feed digestion. Much research has been directed toward methane abatement strategies to be used in ruminants and has been reviewed elsewhere. Abatement strategies are often limited by the diet fed, the management conditions, physiological state and use of the animal, as well as government regulations; resulting in difficulties applying a one size fits all approach to the problem of enteric methane mitigation. To this end, the aim of this paper is to provide background on enteric fermentation and methanogens, as well as some of the methane abatement strategies that have shown efficacy in vivo.

Methane and Ruminants
Greenhouse gases such as carbon dioxide, methane, nitrous oxide, and ozone contribute to climate change and global warming through their absorption of infrared radiation in the atmosphere. Methane is classified as a trace gas and is estimated to have a total global concentration of 1774 ± 1.8 parts per billion (ppb), with a total increase of 11 ppb since 1998 [9]. Methane is an especially potent trace gas due to its global warming potential, 25 times that of carbon dioxide, and its 12-year atmospheric lifetime; it is the second largest anthropogenic greenhouse gas, behind carbon dioxide . Also, methane is able to increase ozone in the tropospheric region of the atmosphere where the greenhouse effect occurs, and increase stratospheric water vapour, both of which can add to the radiative force of the gas by approximately 70%. Globally, 50–60% of methane emissions are from the agricultural sector, specifically from livestock production operations; the principal source of methane is from ruminant animals .
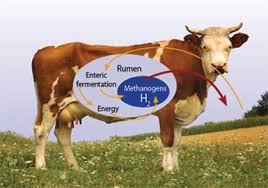
Domesticated ruminants, such as cattle, sheep, and goats produce as much as 86 million metric tonnes (Tg) of methane per year . Approximately 18.9 Tg are from dairy cattle, 55.9 Tg are from beef cattle, and 9.5 Tg are from sheep and goats . Data from Johnson and Ward, estimates the global yearly methane contribution of buffalo to be 6.2–8.1 Tg, 0.9–1.1 Tg from camels, and methane production within the hindgut of pigs and horses to be approximately 0.9–1.0 Tg and 1.7 Tg, respectively.
Methane is produced in the rumen as a product of normal fermentation of feedstuffs. Although methane production can also occur in the lower gastrointestinal tract, as in nonruminants, 89% of methane emitted from ruminants is produced in the rumen and exhaled through the mouth and nose . As methane is exhaled into the atmosphere, the ruminant suffers a loss of ingested feed-derived energy of approximately 2–12%, depending upon the diet .
The loss of methane to the atmosphere varies based on the ruminant species. Estimates of diet-derived energy losses from methane for dairy cattle, range-cattle, and feedlot cattle vary from 5.5–9.0%, 6.0–7.5%, and 3.5–6.5%, respectively . For buffalo and camels, a loss of diet energy in the form of methane ranges from 7.5–9.0% and 7.0–9.0%, respectively . Estimates of methane losses from ruminants also vary based on geographical location, feed quality, feed intake, feed composition, and the processing of the feed .
Methanogens
Characteristics and Ecosystem
Methanogens belong to the domain Archaea and the phylum Euryarchaeota [16]. Unlike Bacteria, methanogens lack peptidoglycan in the cell wall, replaced by pseudomurein in Methanobrevibacter and Methanobacterium, heteropolysaccharide in Methanosarcina, and protein in Methanomicrobium . All methanogens have coenzyme F420, which is a cofactor necessary for enzymes such as hydrogenase and formate dehydrogenase, and received its name due to its absorbance at 420 nm, which allows it to fluoresce blue-green at 470 nm . Another coenzyme characteristic of methanogens is coenzyme M, which is either produced by the methanogens, such as Methanobacterium, or is required from an external source, which is the case for Methanobrevibacter ruminantium . Coenzyme M, or 2-mercaptoethanesulfonic acid, is methylated to produce methane .
The cell characteristics can vary greatly among methanogens as well. Methanobrevibacter ruminantium is rod shaped with variable motility and is able to use hydrogen and carbon dioxide, and formate as substrates for methane production . Methanobacterium formicicum, which is in the same order (Methanobacteriales) as Methanobrevibacter, can be rod or filament shaped without motility, and is able to use the same substrates as Methanobrevibacter . Methanomicrobium mobile is rod shaped and is motile, using both hydrogen and carbon dioxide, as well as formate to produce methane. Finally, Methanosarcina barkeri and Methanosarcina mazeii are both coccoid shaped, but without motility . The order Methanosarcinales contains the only methanogens with cytochromes, and can grow on the broadest range of substrates . Cytochromes are membrane-bound electron carriers that play a role in the oxidation of methyl groups to carbon dioxide. Methanosarcina barkeri is able to produce methane from hydrogen and carbon dioxide, acetate, methylamines, and methanol, whereas Methanosarcina mazeii can use the same substrates except hydrogen and carbon dioxide .
Methanogens with cytochromes have a growth yield of 7 g per mole of methane on hydrogen and carbon dioxide, and have a doubling time of greater than 10 hours. Methanogens without cytochromes have a growth yield of 3 g per mole of methane on hydrogen and carbon dioxide, and have a doubling time minimum of 1 hour . More in-depth discussion of taxonomy can be found in Garcia et al. and a review of methanogens with emphasis on cytochromes can be found in Thauer et al.
Methanogens are not only confined to the rumen in cattle and other ruminants. There has been recent interest in the presence of methanogens in the intestine of humans and Archaea have been found using 454 pyrosequencing in higher abundance in the large intestine of obese individuals . Real-time PCR has been used to detect Methanobrevibacter smithii and Methanosphaera stadtmanae from human feces , and methanogens Methanobrevibacter gottschalkii, Methanobrevibacter thaueri, Methanobrevibacter woesei and Methanobrevibacter wolinii have been cultured from the feces of horse, cow, goose, and sheep, respectively. Finally, Methanobrevibacter oralis was isolated in subgingival sites of patients with periodontal disease .
Methane Production
The principal methanogens in the bovine rumen utilize hydrogen and carbon dioxide, but there is a group of methanogens of the genus Methanosarcina that grow slowly on hydrogen and carbon dioxide and therefore maintain a distinct niche by utilizing methanol and methylamines to produce methane. Formate, which is formed in the production of acetate, can also be used as a substrate for methanogenesis, although it is often converted quickly to hydrogen and carbon dioxide instead. Volatile fatty acids (VFA) are not commonly used as substrates for methanogenesis as their conversion into carbon dioxide and hydrogen is a lengthy process, which is inhibited by rumen turnover . Therefore, methanogenesis often uses the hydrogen and carbon dioxide produced by carbohydrate fermentation, as VFAs are formed. By removing hydrogen from the ruminal environment as a terminal step of carbohydrate fermentation, methanogens allow the microorganisms involved in fermentation to function optimally and support the complete oxidation of substrates. The fermentation of carbohydrates results in the production of hydrogen and if this end product is not removed, it can inhibit metabolism of rumen microorganisms .
The principal methanogens in the bovine rumen utilize hydrogen and carbon dioxide, but there is a group of methanogens of the genus Methanosarcina that grow slowly on hydrogen and carbon dioxide and therefore maintain a distinct niche by utilizing methanol and methylamines to produce methane. Formate, which is formed in the production of acetate, can also be used as a substrate for methanogenesis, although it is often converted quickly to hydrogen and carbon dioxide instead. Volatile fatty acids (VFA) are not commonly used as substrates for methanogenesis as their conversion into carbon dioxide and hydrogen is a lengthy process, which is inhibited by rumen turnover . Therefore, methanogenesis often uses the hydrogen and carbon dioxide produced by carbohydrate fermentation, as VFAs are formed. By removing hydrogen from the ruminal environment as a terminal step of carbohydrate fermentation, methanogens allow the microorganisms involved in fermentation to function optimally and support the complete oxidation of substrates. The fermentation of carbohydrates results in the production of hydrogen and if this end product is not removed, it can inhibit metabolism of rumen microorganisms .
Relationship with Other Microorganisms
Methanogens are known to have symbiotic relationships involving interspecies hydrogen transfer with rumen microorganisms, especially with rumen protozoa where the methanogens can be associated intracellularly and extracellularly [30]. Common protozoa in the bovine rumen found to have such a relationship are from the genera Entodinium, Polyplastron, Epidinium, and Ophryoscolex, while the methanogens most often associated with protozoa are from the orders Methanobacteriales and Methanomicrobiales [30]. Anaerobic fungi, such as Neocallimastix frontalis, have also been found to have a relationship with methanogens involving interspecies hydrogen transfer whereby the fungi’s enzymatic activity has increased and metabolism has shifted towards acetate production .


No comments:
Post a Comment